脳の信号伝達を支える糖鎖を発見
糖鎖合成酵素MGAT5Bが伝達の速度とタイミングを制御
2026年7月14日
岐阜大学
日本医科大学
脳の信号伝達を支える糖鎖を発見 〜糖鎖合成酵素MGAT5Bが伝達の速度とタイミングを制御〜
本研究のポイント
・MGAT5Bは、O-マンノース(Man)型糖鎖と呼ばれる糖鎖の枝分かれを作る酵素で、脳特異的に存在し、脱髄(だつずい)疾患や脳腫瘍などに関わることがわかっています。
・MGAT5Bを欠損したマウスでは、神経の効率的な信号伝達に必要なランビエ絞輪という構造が異常に広がり、神経伝達が遅くなるだけでなく、信号が伝わるタイミングにばらつきが生じることで、運動能力が低下していました。
・MGAT5Bは、ランビエ絞輪の形成に必要なNF186というタンパク質に付いているO-Man型糖鎖を修飾することで、NF186の機能を調節していることがわかりました。
研究概要
岐阜大学糖鎖生命コア研究所の木塚 康彦教授、自然科学技術研究科修士課程2年生の冨田 崇羽さん、日本医科大学大学院の加藤 大輔大学院教授らの研究グループは、広島大学、岐阜薬科大学、東北医科薬科大学との共同研究で、脳においてO-マンノース(Man)型と呼ばれる糖鎖が神経の速い伝達に不可欠であることを発見しました。
タンパク質に付く糖鎖には膨大な種類が存在しており、その形はタンパク質によって異なります。これら糖鎖は、細胞の中で多くの糖鎖合成酵素の働きによって作られ、様々な生命現象や疾患との関わりが報告されています。
本研究では、脳において脱髄疾患などに関わるO-Man型糖鎖に着目しました。O-Man型糖鎖は、脳に特異的に存在するMGAT5B(別名GnT-IX)と呼ばれる糖鎖合成酵素によって枝分かれ構造が作られます。MGAT5Bは脱髄疾患や脳腫瘍との関わりが報告されていますが、正常な脳における役割はよくわかっていませんでした。
本研究では、MGAT5Bを欠損するマウスを用いて解析を行いました。その結果、MGAT5B欠損マウスでは、ランビエ絞輪とよばれる神経の構造が異常に広がり、それにより神経伝達が遅くなるだけでなく、信号が伝わるタイミングにばらつきが生じ、協調運動能力が低下することなどが明らかになりました。本研究は、脳における糖鎖の新しい機能を明らかにしたもので、脱髄疾患などの病態解明にも繋がることが期待されます。
本研究成果は、現地時間2026年7月13日にCommunications Biology誌のオンライン版で発表されました。
なお、本研究は、科学技術振興機構(JST)の創発的研究支援事業の支援を受けて実施しました。
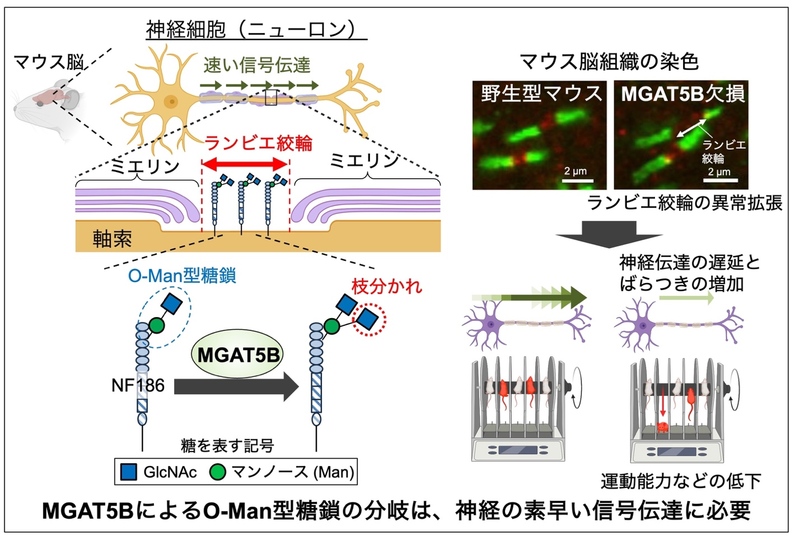
本研究の概要図 (図の作製には一部BioRenderを用いた)
研究背景
糖鎖 1)とは、グルコースなどの糖(動物では約10種類の糖が存在)が枝分かれしながら鎖状につながったもので、多くはタンパク質や脂質などに結合した状態で存在しています。タンパク質に付いている糖鎖には様々な形のものがあり、タンパク質ごとに糖鎖の形が異なること、また同じタンパク質でも、臓器ごとに糖鎖が異なること、健康なときと病気のときとで糖鎖の形が変化することなどが知られています。糖鎖の形が変わることで、タンパク質の機能が変化したり、病気の発症や進行の程度が変化することから、糖鎖の変化や働きの仕組みを理解することは、生命や病気の原因を理解するために重要です。
タンパク質に付く糖鎖は、細胞の中で糖転移酵素 2)(糖鎖合成酵素)と呼ばれる酵素の働きによって作られます。ヒトの体内には、約180種類の糖転移酵素が存在し、それらの働きが厳密に制御されることで膨大な種類の糖鎖が作られます。
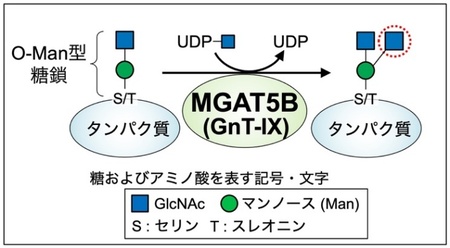
図1 MGAT5Bが作る糖鎖構造
O-Man型糖鎖は、タンパク質のセリン(S)またはスレオニン (T)に付く糖鎖で、MGAT5Bによって枝分かれ構造が作られる。
MGAT5B (GnT-IXとも呼ばれる) 3)は、脳に特異的に存在する糖転移酵素で、タンパク質に付いた O-マンノース(Man)型糖鎖 4)と呼ばれる糖鎖に作用して、枝分かれ構造を作る酵素です(図1)。これまでの研究で、MGAT5Bを欠損したマウスでは脱髄 5)疾患からの回復が早いことや、MGAT5Bを欠損させたがん細胞では、脳腫瘍の一種であるグリオーマ 6)の生育が抑えられることなどが報告され、MGAT5Bが作る枝分かれしたO-Man型糖鎖と疾患との関わりが明らかになってきています。このように、MGAT5Bと疾患との関わりがわかってきた一方で、生理的な状況の脳においてMGAT5Bが果たす役割についてはよくわかっていません。MGAT5Bは、脳を構成する複数の細胞の中でも特に神経細胞(ニューロン)に多く存在していることから、枝分かれしたO-Man型糖鎖は神経細胞の機能に関与していると考えられます。そこで本研究では、MGAT5B欠損(KO 7)) マウスを用い、神経細胞の機能に着目して本糖鎖の役割について調べました。
研究成果
O-Man型糖鎖は、限られた種類のタンパク質にのみ付いていることが知られています。神経細胞に存在するタンパク質では、O-Man型糖鎖を持つものとしてNF186 8)が知られています。またNF186は、神経細胞の中でも、ランビエ絞輪 9)と呼ばれる部分に存在することがわかっています。ランビエ絞輪とは、神経細胞が持つ軸索(長い突起)のうち、絶縁体であるミエリン 10)が巻き付いていない領域のことで、神経細胞はランビエ絞輪ごとに飛び飛びで電気信号を伝達すること(跳躍伝導)で、速い信号伝達を可能にしています。NF186を欠損するマウスでは、ランビエ絞輪が形成されなくなることから、NF186はランビエ絞輪の形成に不可欠であることが知られています。そこで本研究では、MGAT5Bが作るO-Man型糖鎖がNF186の働きやランビエ絞輪の形成に関わる可能性について調べました。
まず、マウスの脳内の複数箇所でランビエ絞輪の染色を行ったところ、MGAT5B KO マウスの脳では、いずれの場所においてもランビエ絞輪の幅が異常に拡張してい ることがわかりました (図2) 。
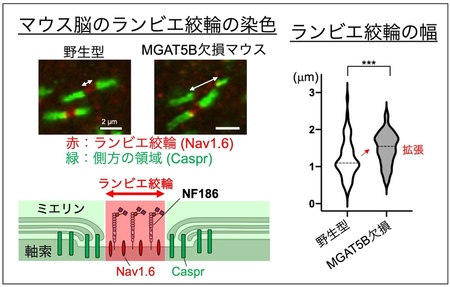
図2 MGAT5B KOマウスにおけるランビエ絞輪の異常拡張
マウス脳内のランビエ絞輪を染色し、幅を測定した。ランビエ絞輪に存在するNav1.6、その側方領域に存在するCasprを染色することでランビエ絞輪を可視化した。右のグラフの点線は中央値を、アスタリスクは野生型とMGAT5B KOの間で統計学的に有意に差があることを表す。
ランビエ絞輪の幅が一定以上になると、幅が大きくなるにつれて神経細胞の伝達速度が遅くなることが知られています。神経の伝達速度の遅延は、運動や認知、精神など脳の様々な機能の低下を引き起こす可能性があります。実際に、MGAT5B KOマウスの運動能力をロタロッド 11)と呼ばれる装置を用いて調べたところ、MGAT5B KOマウスでは、運動能力が低下していることがわかりました(図3A)。そこで次に、脳内 で運動を司る領域(運動野 12))に存在する神経の伝達速度を野生型とMGAT5B KOマウスで比較しました。その方法として、神経細胞の末端を光で刺激し、その信号が軸索を伝わる速度を電極によって測定しました (図3B左)。その結果、MGAT5B KOマウスでは、野生型マウスと比べて、運動野の神経伝達速度 が遅くなるだけでなく、信号が伝わるタイミングにばらつきが生じることがわかりました(図3B右)。これらのことから、MGAT5Bが作るO-Man型糖鎖の枝分かれは、運動野などの神経において、ランビエ絞輪の形成と電気信号の伝達機能に極めて重要であることがわかりました。
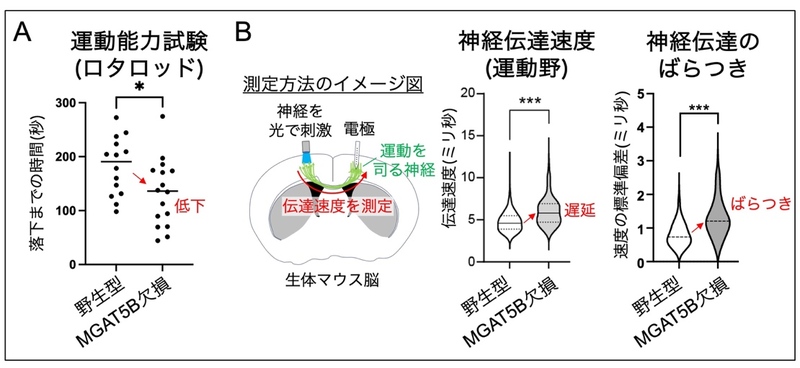
図3 MGAT5B KOマウスの運動能力異常と神経伝達速度の低下
A: ロタロッドと呼ばれる装置を用いた運動試験の成績。回転する棒から落下するまでの時間を計測し、短いほど運動能力が低下することを表す。B: 脳の運動野における神経伝達速度の測定。光によって電気信号を発するように遺伝子操作した神経の末端を光で刺激した。神経の軸索を電気が伝わる速度を電極により計測し、速度とばらつきを表示した。
最後に、O-Man型糖鎖の変化がなぜランビエ絞輪の拡大を引き起こしたのか、その仕組みを調べました。枝分かれしたO-Man 型糖鎖が付いているNF186を脳から精製し、結合しているタンパク質を探索したところ、CNTN1 (Contactin 1 13)) と呼ばれる、ランビエ絞輪の境界の形成に不可欠なタンパク質と結合していることがわかりました。これらのことから、NF186の持つO-Man型糖鎖は、CNTN1との結合に関与することで、ランビエ絞輪の形成を調節しているのではないかと仮説を立てました。これを検証するため、MGAT5Bを持たない細胞と多く持つ細胞に、NF186とCNTN1を発現させ、両者の結合の度合いを共沈降実験によって調べました。CNTN1をビーズを用いて沈降させ、共沈降してくるNF186の量を調べたところ、MGAT5Bを多く持つ細胞の場合では、CNTN1に結合して共沈降してくるNF186の量が少ないことがわかりました (図4) 。このことから、MGAT5Bの作るO-Man型糖鎖の枝分かれ構造は、NF186とCNTN1の結合を抑制しており、MGAT5B KOマウスでは、糖鎖が変化して両者の結合が異常に強くなることで、ランビエ絞輪の幅が拡張していると考えられました (図5) 。
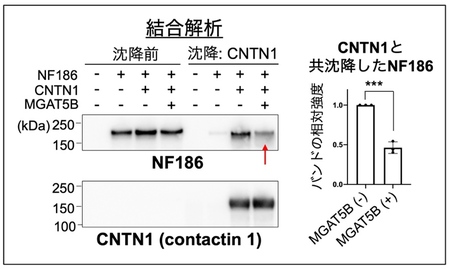
図4 MGAT5BによるNF186とCNTN1の結合の抑制
MGAT5Bを持たない細胞(-)と多く持つ細胞(+)に、NF186とCNTN1を発現させた。CNTN1をビーズにより沈降させ、CNTN1と結合して共沈降してくるNF186の量を、ウエスタンブロッティングと呼ばれる方法で調べた。バンドが濃いほど結合が強いことを表し、赤矢印は結合が弱くなったことを表す。右側は、共沈降したNF186のバンドの強さを表したグラフ。
今後の展開
本研究により、MGAT5Bが作るO-Man型糖鎖の枝分かれ構造は、NF186とCNTN1との結合を抑制することで、神経細胞のランビエ絞輪の形成、神経の電気信号の伝達、ひいては個体の運動機能の調節に重要な役割を果たすことがわかりました。本成果は、脳における糖鎖の新たな役割を明らかにしたものであり、今後、どの糖鎖がどのように脳機能へ関与しているのかを明らかにする手がかりの一つになると期待されます。さらに、MGAT5Bは脱髄疾患やグリオーマに関与していることから、本研究の成果は今後、これら神経系疾患の病態解明に貢献することが期待されます。
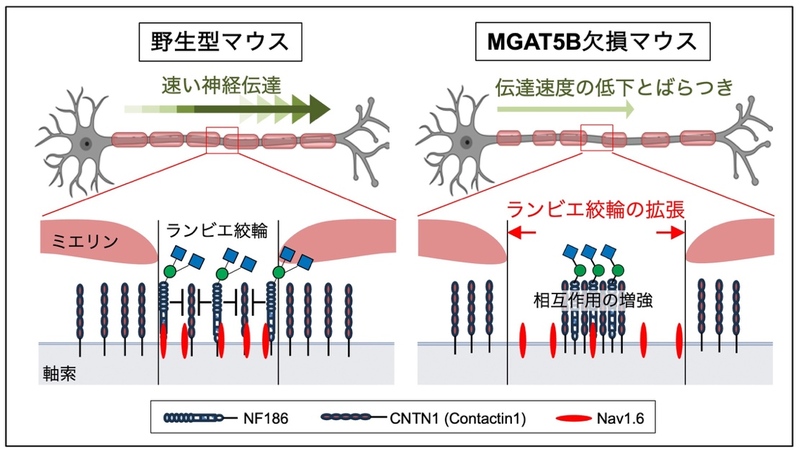
図5 研究結果をまとめたモデル
MGAT5Bが作るO-Man型糖鎖はNF186に付いており、NF186とCNTN1の結合を抑制して適切なランビエ絞輪の幅を形成する。MGAT5B KOマウスでは、NF186とCNTN1との結合が異常に増強され、ランビエ絞輪の末端のCNTN1が減少してランビエ絞輪が広くなると考えられる。図の作製には一部BioRenderを用いた。
【用語解説】
1) 糖鎖:グルコース (ブドウ糖) などの糖が鎖状につながった物質。遊離の状態で存在するものもあれば、タンパク質や脂質に結合した状態のものもある。デンプン、グリコーゲンなどの多糖は数多くの糖がつながり、糖鎖だけで遊離の状態で存在する。一方タンパク質に結合したものは、数個から20個程度の糖がつながったものが多い。
2) 糖転移酵素:糖鎖を合成する酵素のことで、ヒトでは180種類程度存在することが知られている。主に、細胞の中のゴルジ体と呼ばれる小器官に存在している。
3) MGAT5B:糖鎖を合成する糖転移酵素の一種で、細胞の中のゴルジ体に存在し、O-Man型糖鎖の枝分かれ構造を作る。GnT-IXとも呼ばれる。
4) O-マンノース型糖鎖:タンパク質に付く糖鎖の種類の1つで、タンパク質のセリン残基(アミノ酸の1文字表記法でSと表記)もしくはスレオニン残基(1文字表記でT)にマンノース(Man)という糖が結合している。この糖鎖の形成異常は、ある種の筋ジストロフィーの原因となる。MGAT5Bはこの糖鎖の分岐構造を作る。
5) 脱髄:神経細胞は、軸索と呼ばれる長い突起に電気を通すことで、シナプスでつながった別の神経細胞に情報を伝達していく。通常、軸索がミエリン(髄鞘)と呼ばれる絶縁体に覆われて保護されることで正常に電気が伝達されるが、さまざまな原因により髄鞘が破壊されることがあり、それを脱髄という。脱髄が起こると神経の正常な信号伝達ができなくなり、しびれや麻痺を引き起こす。
6) グリオーマ:脳腫瘍の一つで、神経膠腫とも呼ばれる。脳を構成する細胞のうち、グリア細胞が腫瘍化したもの。脳腫瘍の中で最も多い。
7) KO (knockout):特定の遺伝子を改変してその機能を失わせること。現在では、世界中の研究室で、ゲノム編集技術などにより培養細胞や実験モデル動植物などの遺伝子のノックアウトが行われている。
8) NF186:脳に存在するタンパク質の一種で、O-Man型糖鎖が付いている。Neurofascin186ともいう。NF186を欠損するとランビエ絞輪ができなくなることから、ランビエ絞輪の形成において中心的な役割を果たすことがわかっている。
9) ランビエ絞輪:神経の軸索において、絶縁体であるミエリン(髄鞘)が巻き付いていない領域のこと。神経細胞は、ランビエ絞輪ごとに電気信号を飛び飛びに伝達する跳躍伝導を行うことで、素早い信号伝達を行っている。
10) ミエリン:神経細胞の軸索に巻き付いている絶縁体で、髄鞘ともいう。オリゴデンドロサイトと呼ばれる細胞がミエリンを形成する。
11) ロタロッド:マウスやラットなどの運動能力や平衡性を調べるための装置。モーターで回転する棒に動物を乗せ、下に落下するまでの時間を測定できる。
12) 運動野:脳の中で運動を司る神経が存在している領域。大脳皮質の中に存在する。
13) Contactin 1:脳などに存在する接着機能を持ったタンパク質で、ミエリンの形成に重要な役割を果たす。他のタンパク質と結合することで、ランビエ絞輪の側方の領域の末端の構造を維持している。
論文情報
雑誌名:Communications Biology
論文タイトル:Branching of O-mannose glycans regulate node of Ranvier organization and saltatory conduction
著者:Shu Tomita, Taichi Nakaishi, Toshiyuki Ishii, Kazuya Ono, Honoka Fujimori, Misuzu Hashimoto, Shiho Ohno, Yoshiki Yamaguchi, Masamitsu Shimazawa, Miyako Nakano, Daisuke Kato*, Yasuhiko Kizuka* (*共同責任著者)
DOI: 10.1038/s42003-026-10622-0
本プレスリリースは発表元が入力した原稿をそのまま掲載しております。また、プレスリリースへのお問い合わせは発表元に直接お願いいたします。
プレスリリース添付画像
本研究の概要図
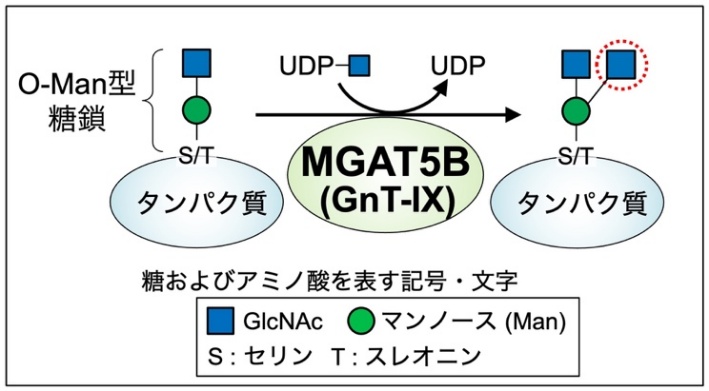
図1 MGAT5Bが作る糖鎖構造
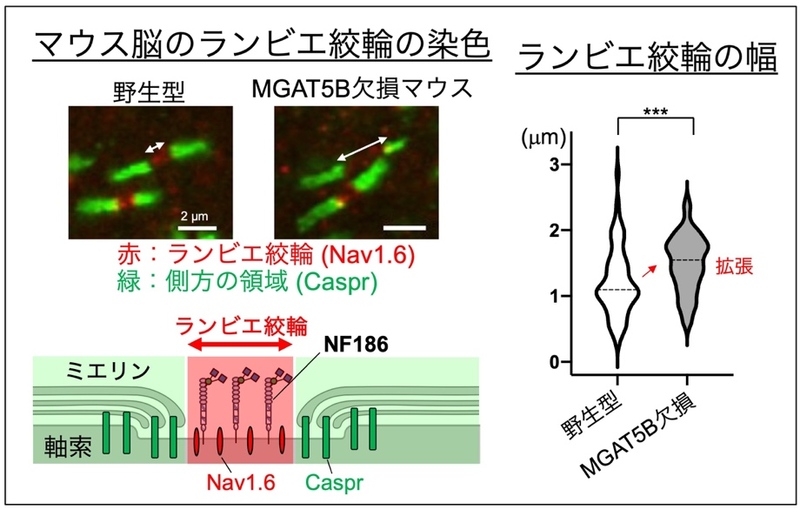
図2 MGAT5B KOマウスにおけるランビエ絞輪の異常拡張
図3 MGAT5B KOマウスの運動能力異常と神経伝達速度の低下
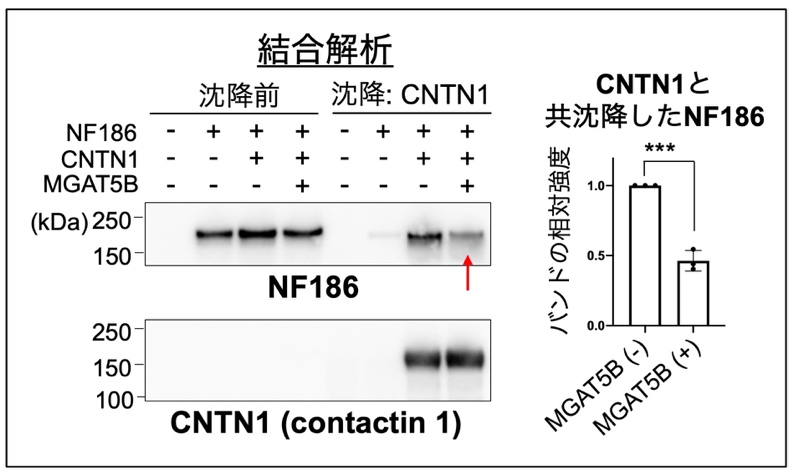
図4 MGAT5BによるNF186とCNTN1の結合の抑制
図5 研究結果をまとめたモデル
このプレスリリースには、報道機関向けの情報があります。
プレス会員登録を行うと、広報担当者の連絡先や、イベント・記者会見の情報など、報道機関だけに公開する情報が閲覧できるようになります。
このプレスリリースを配信した企業・団体
- 名称 国立大学法人東海国立大学機構岐阜大学
- 所在地 岐阜県
- 業種 大学
- URL https://www.gifu-u.ac.jp/

過去に配信したプレスリリース
大質量星団は、銀河最強クラスの宇宙線加速器だった
7/27 15:00

免疫介在性中枢神経疾患の病態メカニズムを解明
7/27 11:00
骨肉腫の「肺転移」を抑制するメカニズムを解明:骨肉腫の新規治療標的を発見
7/16 14:00
脳の信号伝達を支える糖鎖を発見
7/14 14:00
宇宙ジェットが分子雲に衝突する現場を発見
6/26 10:00
乳児の健康を支える「母乳オリゴ糖」をつくる培養細胞を開発
6/25 15:00
温暖化によるアユ生態の南方化
6/15 14:00
女性特有の症状・疾患への初期対応を支援
6/12 14:00
特定の酵素の作用によって「溶ける/固まる」 新しいゼリー素材を開発
6/4 09:00
がん遺伝子RASを標的とするタンパク質型抗がん剤候補を開発
5/29 14:00
12チャンネル3次元音響センサアレイによる非定常騒音の時空間可視化技術を開発
5/28 14:00












